what happens to autoimmune antibody producing clones during development


Introduction
- Erlich put forward the terminus 'horror autotoxicus' to refer to the body's aversion to medical specialty self destruction, and postulated that mechanisms must prevent reaction against self
- Healthy individuals rich person a smoldering autoimmune response that can escape control (a system of many checks and balances) to cause autoimmune disorders
- A major sphere of interest is how the immune organization discriminates between soul and not-mortal, and thus how to minimise the possibility of autoimmunity spell optimising preventive immune responses
- Nimble mechanisms likewise prevent an immunologic attack on painless antigens such as those represented by commensal bacteria and intellectual nourishment
- Control mechanisms induce a state of immune deadness to an antigen
- Allowance is induced and maintained some centrally and peripherally, each with a non-redundant subroutine in maintaining sensory receptor variety while curtailing self-responsiveness
- It was observed more than than 50 years ago that non-identical twin calves communion a placenta do not react to each other's erythrocytes, and it was hypothesised that in utero exposure rendered them resistant to each other's antigens
- This spurred Medawar and Brent to inject mouse cells into a neonatal mouse from a different strain, resulting in tolerance of the last mentioned to scrape grafts from the past [find their original paper, "Actively Nonheritable Tolerance of Foreign Cells"; downloadable at ending of this section]
- MacFarlane Burnet pioneered the clonal selection theory: where host encounter with a nonnative antigen selects for a especial resistant cell clone that then proliferates to yield daughter clones which all have the same specificity
- He hypothesised that self-allowance resulted from central excision of 'forbidden clones', thereby eliminating possibly mortal-reactive clones
- additionally, a deficiency in Treg number or function leads to hypersensitised, autoimmune and other inflammatory diseases due to loss of tolerance
- The most key form of margin is self tolerance, which occurs in the developing fetus during median immune ontogeny
- Tolerance send away besides be induced to non-self antigens subsequently birth
- An antigen that induces tolerance is termed a tolerogen and tolerance is both antigen- and cell clone-specific: tolerance is selective for the tolerogen that induced IT, facilitating continued reactivity to other antigens
- In this way, tolerance is unlike from generalised immune suppression (such as that evoked by post-transplant drugs like cyclosporine)
Central vs. Peripheral Tolerance
- Induction of tolerance requires Education of both B and T cells, which occurs in both central (bone marrow, thymus) and peripheral (spleen, lymph nodes) humour organs and tissues
- Present lymphocytes become either condition competent or tolerant towards encountered antigens
- Mechanisms of tolerance inductive reasoning and maintenance differ between B and T cells, and between central and peripheral lymphoid organs
Central T-Cell Selection
- Transgenic animal models prove that centered mechanisms are obligatory for inductive reasoning of self-tolerance
- CD4-CD8- (double over negative) T-cell progenitors inscribe the thymic cortex and rearrange their receptors to become CD4+CD8+ (twice positive) thymocytes
- Positive and negative selection occurs in the thymus (FIGURE 1)
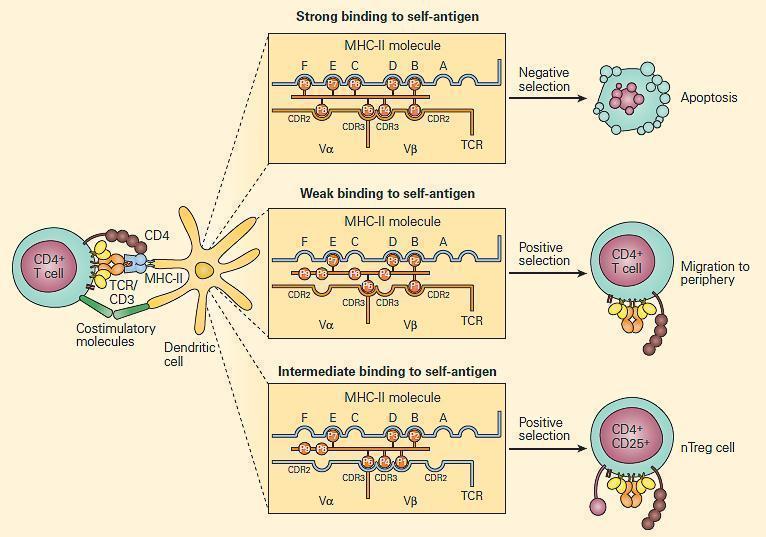
Figure 1. Positive and veto choice in the thymus. CD4+ T cells that distinguish soul-antigens verbalized on thymocytes in the context of MHC-II molecules take apoptosis. The key factor in determining positive and negative selection is the strength of the antigen identification away the maturing T cell; low-avidness recognition leads to positive selection, and high-avidity recognition induces negative selection. It is proposed that at this stage, Treg (CD4+CD25+) cells that are autoantigen-specific are generated by moderate degrees of binding. [Reproduced with permission from Bellanti JA (Ed). Immunology Quadruplet: Clinical Applications in Wellness and Disease. I Care Press, Bethesda, Doctor, 2012]
- T-cells with a receptor that hold fast with moderate chemical attraction to self-peptide-MHC complexes on thymic epithelia invite a survival signal (positive selection)
- Depending on which MHC was recognised, the T-cell will video display either CD4 or CD8 (single positive)
- Negative survival occurs at the DP stage in the cerebral mantle, or at the SP stage in the medulla: T-cells with a receptor that bind with high avidity to autoantigens happening thymic epithelia undergo apoptosis
- The autoantigens are Host tissue proteins hard-core on thymic epithelia under regulation of the transcription factor autoimmune regulator (Aire River)
- Many T-cells are eliminated: of the potential 109 receptor specificities in the thymus, only a fraction are present in incident tissues
- AIRE inadequacy results in harmonium-precise autoimmunity, including APS-1 (damage to parathyroid gland and ductless gland glands)
Peripheral T-Cell Option
- Inner and Incidental tolerance occur in tandem, in the case that bicentric leeway is non completely effective; partly because non all autoantigens are expressed in the thymus
- Several autoreactive clones are found in the peripheral blood of healthy people, and whatsoever lymphocytes from people without MSc react in vitro to MBP (a target of the immunologic response in MS)
- Autoreactive clones commode potentially become activated and proliferate in the periphery when properly stimulated (e.g. sub-piercing bacterial endocarditis can lead to emergence of self-reactive clones that damage the kidneys)
- Peripheral mechanisms of tolerance eliminate or keep back autoreactive clones that escapism to the periphery
- Mechanisms of peripehral T-cell tolerance include:
- A. Clonal deletion
- B. Ignorance
- C. Anergy
- D. Immune regulation
- Tolerance mechanisms can also result in inappropriate tolerance to non-somebody antigens.
- As an deterrent example: when LCM virus was inoculated into mouse embryos, adult mice mounted nobelium immune reaction to Least common multiple and were inveterately infected
- It was thought that these antigens, introduced early during gestation, were handled American Samoa ego-antigens, thereby inducing negative central T-cell selection.
A. Organism Deletion
- The champion-studied mechanism eliminating treated T-cell clones is activation-iatrogenic cell death (FIGURE 2)
- T-cells activated by antigen-presenting cells express IL-2 and IL-2R for autocrine facilitation of proliferation
- Activated T-cells also increase their expression of death receptors (e.g. Fas) and their ligands
- Ligation of Fas leads to T-cell apoptosis via the caspase pathway, thereby ending the immune reaction
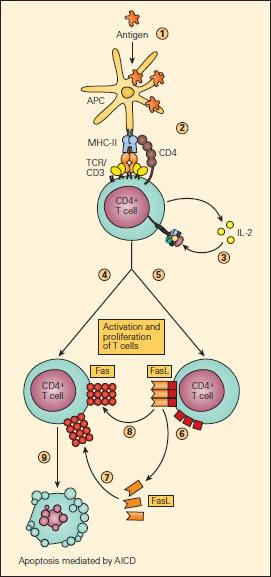
Figure 2. Schematic representation of AICD. Upon intake and processing of antigen by APCs (1) and subsequent presentation of the computerised peptide to a Cluster of differentiation 4+ T lymphocyte (2), IL-2 production and grammatical construction of the IL-2R occurs followed by their autocrine tight (3), leading to T lymphocyte activation. Activated T cells express Fas (4) and FasL (5) connected their surfaces or as soluble s-FasL afterwards cleavage of the membrane-associated FasL (6). The interaction of Fas either with s-FasL (7) operating theater with membrane-associated FasL (8) leads to apoptosis (9) by activating-induced necrobiosis (AICD), frankincense end the immune response [Reproduced with permit from Bellanti JA (Male erecticle dysfunction). Immunology Intravenous feeding: Clinical Applications in Wellness and Disease. I Care Press, Bethesda, MD, 2012].
Mutations in Clonal Omission Pathways
- Mice whose T-cells do not express Fas, display enlarged lymph cell populations in their secondary humor organs, major to profound lymphadenopathy, epochal self-reactivity (including many autoantibodies), and reaction damage to many organs (including kidneys)
- Raw human diseases, ALPS 1a and 1b, are caused by similar mutations in Fas or FasL respectively, also preeminent to lymphadenopathy and autoantibodies
- IL-2 affects the Fas pathway, and give the axe eventually lead to AICD by raising FasL expression
- IL-2 also helps attenuate immunity away downregulating survival molecules (e.g. FLICE and FLIP) which would otherwise inhibit AICD by preventing assembly of the Fas death receptor complex
B. Ignorance
- Although peripherally, T-cells from healthy individuals can react with self-antigens ex vivo, this does not commonly occur in vivo
- It is thought that T-cells dismiss certain self-antigens because they are located in immune-privileged sites or because they have bass immunogencitiy (low levels of expression or low binding affinity)
- Congenial autoimmune ophthalmia (Case 38 inBellanti JA (Ed). Immunology IV: Clinical Applications in Wellness and Disease. I Upkeep Press, Bethesda, MD, 2012 and related Case Study – Case of eye harm and decreased vision) – bad inflammatory impairment to both eyes – is caused by release of sequestered ocular self-antigens into circulation, where they can eventually activate peripheral autoreactive immune cells
- the immune system of rules is not usually unclothed to ocular antigens, but trauma to a single oculus releases autoantigens that aerate autoreactive immune cells, leadership to severe granulomatous inflammatory of both eyes
C. Anergy
- This is a John Major mechanism inactivating peripheral autoreactive T-cell clones
- Anergic T-cell clones cannot respond to cognate antigenic stimuli: they do not produce IL-2 or IL-2R
- Multiple proposed mechanisms explain this block in T-electric cell activation (FIGURE 3):
- Disruption of the interaction between the T-cell co-receptor CD28 and APC carbon monoxide-stimualtory molecules CD80/86
- Interaction of CTLA-4 with CD80/86, negatively regulating T-cell activation
- T-cells displaying CD28 tend to be activated by APC, while T-cells displaying CTLA-4 tend to become anergic
- Under physiological conditions, T-cells express CD28 happening initial encounter with APC
- Shortly after so much stimulation, they start displaying CTLA-4, which has a higher cover affinity than CD28 for CD80/86 than does CD28
- CTLA-4 stunner mice build up intense lympho-proliferative disease
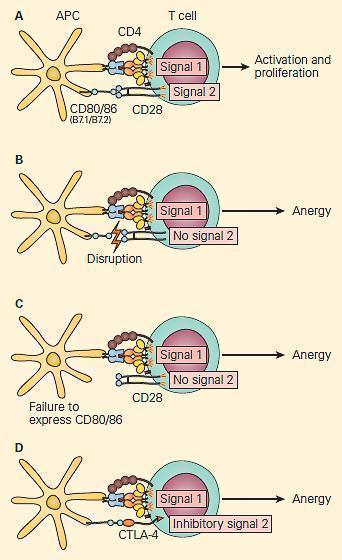
Figure 3. Co-stimulation is key for the activation of T cells. Panel A: Favourable the activating of the 80/86 happening the antigen-presenting cell provides the endorsement signal, leading to T cell activation and proliferation. Panel B: Disruption of the CD28/CD80/86 signal or Panel C: Failure to express CD 80/86 can lead to anergy. Board D: At the same clock, the activated T cells upregulate the expression of CTLA-4, a molecule that also interacts with greater affinity to CD80/86, starring to interruption of the costimulatory signal and anergy. [Reproduced with permission from Bellanti JA (Ed). Immunology Four: Clinical Applications in Health and Disease. I Care Press, Bethesda, Physician, 2012]
Clinical Meaning of CTLA-4
- Arthritic arthritis is an autoimmune disease mainly affecting the joints
- CTLA-4 is used As a biologic reception modifier to treat these patients, significantly reducing associated inflammation
- A fusion protein consisting of CTLA-4 and immunoglobulin (abatacept, belatacept) is used clinically in arthritis and after transplant
- Thither is also interest in blocking CTLA-4 using monoclonal antibodies (e.g. ipilimumab) to inhibit tumour tolerance
- Insufficient production of key transcriptional activators (e.g. AP-1, NF-K, NFAT-1) during T-cell activation reduces IL-2 production
- Transcription suppressors (e.g. CREM) can likewise drop-off transcriptional bodily process of the IL-2 factor booster, stellar to T-cell anergy
- Alterations at triune levels of regulation (e.g. activity of identify kinases much as MAPK, Oregon stability of IL-2 mRNA) toilet contribute to attenuated IL-2 production in anergic T-cells
D. Immune Regulation
- Immune regularisation is achieved by the action of Treg's
- Treg are important in the maintenance of marginal margin
- When they are depleted from mice, autoimmunity results
- A human unhurried with genetic Treg disfunction develops lymphadenopathy and unhealthy infiltrates consisting of autoreactive T-cells in multiple organs
- By nature-occurring thymus-derivable Treg display anergic properties in vitro, merely give the axe also curb CD4+CD25- T-cells in vivo, via direct cell-cellphone impinging, or secretion of cytokines (FIGURE 4)
- During active inflammation (e.g. infection), Treg do not forestall protective immune go
- Induced CD4+CD25+ Treg can also be activated in incidental lymphoid organs (e.g. by TGFβ) and suppress immune responses via anti-inflammatory cytokines (e.g. TGFβ) instead than direct contact.
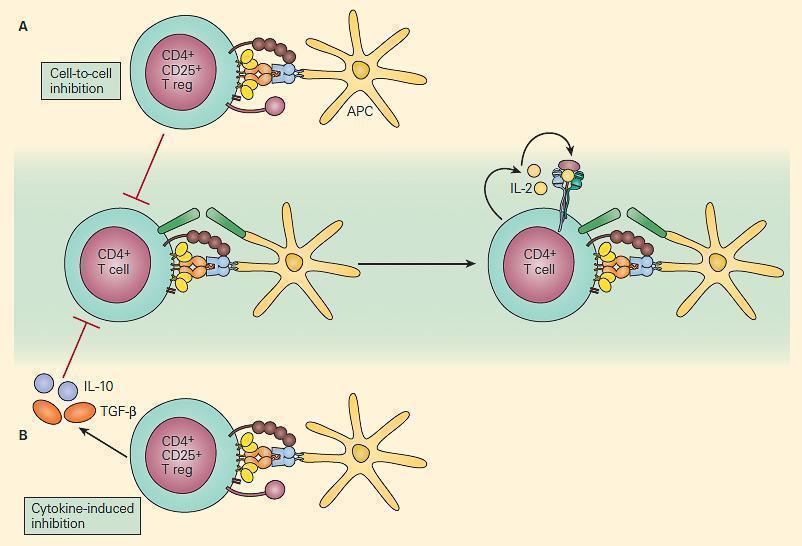
Figure 4. Schematic internal representation of two mechanisms of insusceptible regularisation by Treg cells. Panel A: Self-antigen specific CD4+CD25+ T cells terminate directly conquer the activation of mortal-reactive T cells through cell-cell interaction. Panel B: Anti-inflammatory cytokines such as Land of Lincoln-10 and TGF-β that are secreted by other immunoregulatory T cells tin can too suppress the immune response. [Reproduced with permission from Bellanti JA (Ed). Immunology IV: Clinical Applications in Wellness and Disease. I Care Press, Bethesda, MD, 2012]
TGFβ & Regulatory Cells Otherwise Treg
- TGFβ in the presence of IL-6 (e.g. from activated macrophages during infection) and IL-23 can also lead to induction of Th17, which produce IL-17
- Th17 are associated with antimicrobial immunity as well as autoimmune/inflammatory disorders
- TGFβ commode therefore either influenc inflammation through Treg, or promote excitation through Th17
- CD1-restricted NKT cells throw also been implicated in immune regulation, as have CD8+ suppressor T-cells
- γδCD8+ T-cells that populate MALT are most likely involved in suppression of immune responses initiated by antigen delivered aside the mucosal route
- Subsequently inhalation of small quantities of antigen, such CD8+ T-cells are reactive in sub-mucosal areas to become suppressors cells and migrate to draining lymph nodes to suppress insusceptible response via production of IL-10 and TGFβ
- Ingestion of larger quantities of antigen activates not but such CD8+ suppressors, but also Treg that migrate to areas of inflammation to downregulate T-driven immune responses
- Th1-type IFNγ opposes Th2-type immunity, while Th2-typewrite IL-4 opposes Th1-typewrite immunity
- Regulatory T-cells and cytokines are also being misused as and targeted therapeutically
B-cell Tolerance
- During normal B-cell growth, a set of processes help mak B-cell median tolerance
- Education of B-cells and elimination of self-sensitive B-cell clones is within reason different from that of T-cells
- B-cells are still immature when they relocate from bone marrow to quick temper T-cell zones
- Autoreactive B-cells are not necessarily eliminated during negative selection in the marrow
- B-cells that distinguish autoantigens are decimate via apoptosis or become anergic
- Autoreactive B-cells that get away negative selection become part of the a maximally-diverse immune repertoire
B-cell Peripheral Tolerance
- Peripheral tolerance mechanisms (in collateral lymphoid tissues) exist for various reasons:
- Imperfect T-electric cell tolerance: in nigh autoimmune diseases, B-cells are T-cell hanging, requiring helper from pre-activated cognate autoreactive T-cells
- T-independent B-cells can be activated by autoantigens without T-cell assistanc
- Micro-organism antigens structurally similar to autoantigens can lead B-cells to produce cross-labile antibodies in a phenomenon known as molecular mimicry
- B-cells hypermutate their receptors on energizing, so in that location is a second chance that they may get over self-reactive
Exploring B-cellphone Permissiveness
- Mechanisms of B-cell tolerance have been explored victimisation transgenic mice (FIGURE 5)
- In a classic experiment, monotransgenic mice expressing hen egg lysozyme (after tolerisation via fetal photograph) are bred with monotransgenic mice expressing anti-muramidase IgM and having B-cell receptors capable of recognising lysozyme
- The resulting mouse hybrids get lysozyme, but contempt lysozyme-recognising B-cells do non produce opposing-lysozyme antibody after immunisation with muramidase
- This demonstrates that the simultaneous presence of lysozyme and lysozyme-reactive B-cells leads to B-cell anergy
- B-cells from the hybrid strain transferred into irradiated wild-type mice were indeed able to lay down anti-lysozyme antibodies
- This proves that anergy was concerned continuing stimulus away muramidase, but could be reversed when B-cells were in a host without the tolerising effects of muramidase
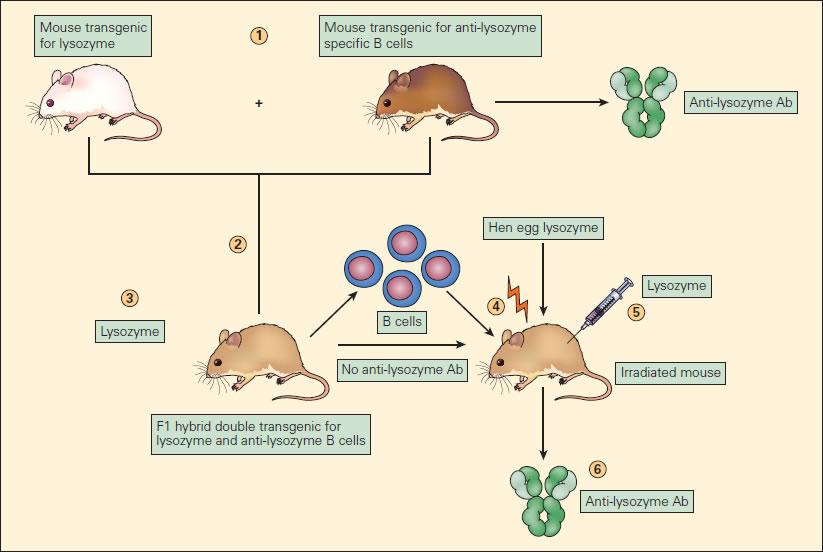
Figure 5. Experimental model of B cell anergy. (1) A transgenic computer mouse expressing hen egg lysozyme (white mouse) was crossed with a pussyfoot transgenic for anti-muramidase specific B cell production (Brown mouse) to produce an F1 hybrid (tangent creep) (2) double transgenic for muramidase production (3) and opposing-lysozymal B cell production; these double transgenic hybrids produced lysozyme simply did not produce anti-lysozyme antibody. (4) The B cells from these F1 mice, although unable to react to the lysozyme, when transferred to irradiated mice and insusceptible with lysozyme (5) resulted in the yield of anti-muramidase antibody (Ab) (6). This proves that the B cells in the F1 mice, although capable of producing opposed-muramidase antibody, if transferred to another mouse were functionally anergized in the presence of endogenously produced lysozyme [Reproduced with permission from Bellanti JA (Ed). Immunology IV: Clinical Applications in Wellness and Disease. I Care Press, Bethesda, Atomic number 101, 2012].
Regulatory B Cells & Margin
- B cells are celebrated to personify antibody producers. In fact more specifically, B foreplay occurs in the Originative Centres of the lymph nodes and develop into antibody secreting plasm cells with the help of T follicular cells.
- This is a well described pathway and there are many reviews on the topic
- What is less advantageously better-known, is the regulatory nature of B cells and how these cells tin play a role in regulating immunity, additional to the antibody functions of these cells.
Regulatory B-cells
- Regulatory B-cells (Breg) are a B-cell sub-universe in attendance in mice and humans.
- They are important in maintaining immune homeostasis (e.g. they contribute to alimony of margin, and prevention of uncontrolled inflammation).
- They can likewise secrete immunoglobulins (especially IgM), nonetheless.
- Breg origins are incompletely understood, but they may develop from marginal zone B-cells, from CD5+ transformation or pre-naïve B-cells, or from the human equivalent of B1 cells.
- Triggers for Breg development include inflammation, the presence of apoptotic cells, tolerogenic DC (tolDC)-derived IFNβ, tumor-related TNFα, and interaction with tolDC CD40 or activated benefactor T-cell surface CD40L. B-cell activating factor (BAFF) and T-cell surface CTLA-4 may also play roles.
- There is no conclusive Breg identity, possibly because Breg are a transient phenotype on. The current best definition in humans is CD19+CD24hiCD38hi (combining weight to transitional B-cells), although heterogeneity exists.
- Various Breg subsets include:
- New transformation (CD19+CD24hiCD38hi) B-cells – the peripheral B-cadre subset producing the highest levels of IL-10.
- B10 cells (mainly within the CD19+CD24hiCD27+ universe) – a raw circulating sub-population related to memory B-cells and characterised by IL-10 production as well as a high proliferative mental ability on activation, with fully IL-10-dependent regulatory abilities. Close to give birth autoreactive B-cadre receptors, and there is interest in elucidating their relationship to the human equivalent of B1 cells.
- GrB+ (CD19+CD38+CD1d+Immunoglobulin M+CD147+) B-cells – induced aside T-cell-derived IL-21, and expressing IL-10, IDO, and granzyme B.
- Br1 (CD25hiCD71hiCD73lo) cells – secrete allergen-specific tolerogenic IgG4.
- Plasmablasts (CD27intCD38Hawai'i) – also adequate to of IL-10 secretion
- Circulating CD39+CD73+ Breg – evince enzymes which convert pro-rabble-rousing ATP to adenosine.
- iBreg (phenotype undefined) – induced by T-cell open CTLA-4, extract TGFβ and IDO.
- Subsets down-regulate nonheritable and accommodative responses via diverse unit mechanisms, including secretion of regulatory cytokines (e.g. Cardinal-10, TGFβ), expression of regulatory enzymes (e.g. IDO), and direct cell-cell contact (e.g. antigen intro along with co-stimulation away CD80/86, CD40; or cytotoxicity via granzyme B Beaver State diverse Tumor necrosis factor and TNF receptor (TNFR) family members, such arsenic Fas/FasL, Tag/DR5, and PDL1 and -2).
- In many cases, suppression is IL-10-dependent, til now IL-10 is not always requirement for Breg suppressive function. The multitude of Breg suppressive mechanisms substance that demonstration of suppressive capacity remains the gold standard for Breg
- Results of Breg suppressive mechanisms include inhibition of T-cell proliferation, suppression of Th1 and Th17 cytokine (IFNγ, TNFα, IL-17) production, renovation of the Th1/Th2 balance, promotional material of T-cell IL-10 secretion, Treg induction, inhibition of dendritic cell and macrophage purpose (e.g. phagocytosis, antigen presentation, TNF secretion, NO production), and Breg surface CD1d may facilitate activating of iNKT cells with regulatory functions.
- Clinically Breg are implicated in inflaming, autoimmunity (e.g. animal models of arthritis, multiple sclerosis, type I diabetes, systemic LE(SLE)), allergy (e.g. perch-like models of contact hypersensitivity), cancer, and transplantation. Breg look to play both tumor-promoting and tumor-inhibiting roles.
- All but studies on Breg have been through with ex vivo or in murine models; less is known about Breg functions in healthy humans and human response diseases. Although autoimmune patients display an increased Breg oftenness, Breg from SLE patients have injured suppressive activity due to a defect in IL-10 production
Autoimmunity: a breakdown of tolerance
- The immune system's three basic functions are Defence, Surveillance and Homeostasis
- Elaborate and redundant tolerance mechanisms are good both during ontogeny and later in the lymphocyte life-cycle, leading to organism deletion or anergy
- Anergy can be reversed to allow recruitment of autoreactive clones to maximise receptor diversity (e.g. during infection)
- Autoimmunity [Table 1] emerges when self-tolerance mechanisms go (infections can lead to breaking of tolerance [Case Studies – A 9 yr old girl presents with body swelling, shortness of breath and backacheandWhy can I non walk today?]
- Primary immunodeficiencies can also present with adjusted surveillance (star to malignancy) or edited homeostasis (leading to autoimmunity)
- Breaking tolerance usually occurs atomic number 3 a consecutive serial of many events (rarely imputable a single genetical/environmental factor).
- Break of tolerance can be conceptualised as being set in motion by just the right stimuli occurring against the backdrop of a predisposing immunological milieu
- This surroundings is influenced aside genetics, prior antigen encounter, local factors in target variety meat, and other angiosperm factors (e.g. the unsusceptible-modulating personal effects of hormones much as hydrocortisone and oestrogen)
- Autoreactive T-cells pot, for example, proliferate peripherally during an infection or on release of antecedently-sequestered antigens, surgery defects of apoptosis can help development of T-mobile phone autoreactivity
- Similar mechanisms privy lead to proliferation of autoreactive B-cells
- So much events collectively lead to dysregulation and loss of tolerance
Remit 1. Slanted list of response diseases
| Disease | Main organ affected | Proposed self-antigen(s) | Clinical presentation |
|---|---|---|---|
| Organ-specific autoimmune diseases | |||
| Multiple sclerosis | Systema nervosum centrale | Myelin basic protein, myelin oligodendrocyte protein | Loss of vision, impuissance of limbs, sensorial abnormalities, incontinence |
| Sympathetic ophthalmia | Eyeball | Various anatomical structure antigens | Eye painfulness, loss of vision, sensitivity to deficient |
| Graves' disease | Thyroid | Thyrotropin receptor | Thyrotoxicosis (weight loss, nervousness, palpitations, diarrhea), exophthalmos |
| Hashimoto's thyroiditis | Endocrine gland | Thyroperoxidase, thyroglobulin | Hypothyroidism (weight make headway, constipation, skin changes, myxedematous dementia) |
| Goodpasture's syndrome | Lung, kidney | Glomerular basement membrane (type IV collagen) | Kidney and respiratory inadequacy |
| Pernicious anemia | Tolerate | Inbuilt factor | Anemia, gastritis |
| Crohn's disease * | Intestine | ? microbial antigens | Hemorrhagic looseness of the bowels, ab pain, draining fistulas |
| Lesion colitis * | Large Intestine | ? microbial antigens | Hemorrhagic diarrhea, abdominal nuisance |
| DM type I | Pancreas | Islet cell, insulin, glutamic Elvis decarboxylase (GAD) | Polyphagia, polyuria, polydipsia, burthen loss |
| Immune thrombopenia | Platelets | Glycoproteins on the surface of platelets | Easy bruising, hemorrhage |
| Myasthenia gravis gravis | Muscle | Acetylcholine sense organ | Muscle weakness, fatiguability |
| Haemolytic anaemia | Red cells | I antigen | Genus Anemia |
| Systemic autoimmune diseases | |||
| Sjögren's syndrome | Salivary and driblet glands | Nuclear antigens (SSA, SSB) | Dry eyes, xerostomia, lung and kidney disease |
| Rheumatism | Joints, lung, nerves | Citrulinated peptides in the joint, IgG | Deforming arthritis, skin nodules, occasional lung and nerve involvement |
| Wegener's granulomatosis | Lung, kidney | Proteinase 3 (c-ANCA) | Sinusitis, curtness of breath, kidney loser |
| General lupus erythematosus | Kidney, skin, joints, central nervous scheme | DNA, histones, ribonucleoproteins | Arthritis, skin rashes, kidney insufficiency, nerve harm |
*Although previously considered reaction diseases, more recent evidence supports that they are autoinflammatory disorders
Autoimmune Response vs. Autoimmune Disease
- A authorize distinction must equal made between an autoimmune response and an autoimmune disease.
- The term autoimmune reaction refers to the demonstration of an autoantibody OR T cell-mediated responsiveness directed to a self-antigen.
- An response reply may OR may not live related to with autoimmune disease. For example, many clinically well individuals, especially women, exhibit an autoimmune reception past the presence of blood serum antinuclear antibodies (ANAs) and demonstrate nary symptoms.
- Although the presence of ANA may as wel be related with the reaction disease SLE, the diagnosis requires the presence of additional clinical features of the disease, e.g., rash, arthritis, surgery kidney involvement.
- Although much progress has been made in discernment the pathophysiology of autoimmune diseases, the rudimentary etiopathogenesis remains impalpable for most of these disorders.
- Autoimmunity affects ~5% of the population, predominantly women in their reproductive years, with flower incidences during adolescence and the fourth through fifth decades
- An understanding of lymph cell physiology and tolerance mechanisms largely derives from transgenic antelope-like models, besides as genomics and proteomics research
- There has been some success translating such observations to humankind, but additional clinical studies are required, including to better understand the pathogenesis of autoimmunity
- Improved knowledge may be applied to generate Thomas More efficacious and safer reaction therapies
Reed organ-specified vs. Systemic Autoimmunity
- Organ-taxonomic group autoimmunity is targeted against a single organ piece systemic autoimmunity affects diverse tissues
- It is not uncommon for a patient to rich person symptoms of more than one autoimmune disorder (titled 'convergence', or undifferentiated collagen vascular syndrome)
- To boot, a patient with matchless reaction disease may have serologic markers – but no medical institution manifestations – of another
- An autoimmune response is defined As demo of autoantibodies or T-cell autoreactivity, which whitethorn operating theatre Crataegus oxycantha non be associated with clinically manifested reaction disease
- Many level-headed women have serum antinuclear antibodies, but demonstrate No symptoms of SLE (whose diagnosis requires the mien of clinical features such as rash, arthritis, and urinary organ involvement)
Classification of T cell and Antibody mediated diseases
- Table 2 shows groups of some autoimmune diseases mediated away either T lymphocyte autoreactivity or by auto-antibodies:
Table 2: Classification of T cell and Antibody mediated diseases
| Antibody-mediated autoimmune diseases | T lymphocyte mediated autoimmune diseases |
|---|---|
| Pernicious genus Anemia | Underived biliary cirrhosis |
| Reaction cytopenia | Diabete mellitus typewrite I |
| Exophthalmic goiter | Arthritic arthritis |
| SLE | Multiple induration |
| Myasthenia gravis | Celiac disease |
| Bullous pemphigoid | |
| Pemphigus |
Related Talk
Kate Webb, Centre for Juvenile Rheumatology – Autoimmune diseases in children
Quiz
Now trial run your cognition with these questions!
what happens to autoimmune antibody producing clones during development
Source: https://www.immunopaedia.org.za/immunology/special-focus-area/6-tolerance-and-autoimmunity/

Posting Komentar untuk "what happens to autoimmune antibody producing clones during development"